< krisa >
<previous> <на главную> <next>
|
|
|
< krisa > <previous> <на главную> <next> |
|
<01> Механизм мышечного сокращения: |
|
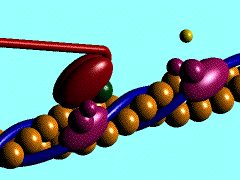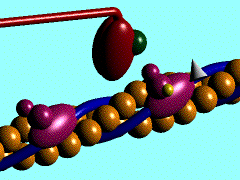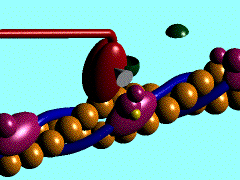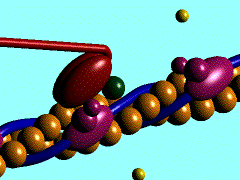
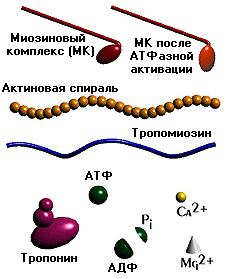
В основе сокращения мышцы лежит укорочение отдельных её клеток - миоцитов [видеозапись сокращений изолированных мышечных клеток, 503 Kb]. Миоциты, в свою очередь, сокращаются в результате укорочения саркомеров миофибрилл (внутриклеточных нитеподобных белковых структур), которое обусловлено скольжением тонких актиновых филаментов вдоль толстых миозиновых по направлению к центру саркомера. Ниже показан механизм такого скольжения - специфический контакт одной из множества "головок" миозина с актиновой нитью и последующий "гребок". В результате одного такого движения саркомер укорачивается лишь на 20 нм (т.е. на 1% своей длины). Поэтому для достижения эффекта движения на макроуровне (например, поднятия предплечья при сгибании руки) необходима целая серия последовательных "гребков", как это показано на схеме. |
|
|
|
|
Схема, демонстрирующая молекулярные механизмы мышечного сокращения с точки зрения теории "скользящих нитей": показан контакт головки миозина с актином (образование "поперечного мостика") и "гребок", продвигающий актиновую нить к середине саркомера (Источник: San Diego State University College of Sciences [www.sci.sdsu.edu] ). Миозиновые нити несут
множество таких "головок", которые
представляют собой биполярно
расположенные выступы длиной около 20 нм,
состоящие примерно из 150 молекул миозина.
Как видно из рисунка, в данной реакции
кроме актина и миозина принимают
участие ещё несколько компонентов. В
первую очередь это тропомиозиновые
нити, лежащие в желобках, образованных
спирально скрученной парой актиновых
нитей. Считается, что в состоянии покоя
тропомиозин расположен так, что
препятствует контакту головки миозина с
актином. Для того, чтобы в случае
необходимости "открыть доступ",
существует так называемый "кальциевый
переключатель": через регулярные
промежутки (около 40 нм) актиновые нити
несут на себе молекулы тропонина.
При связывании с Са(2+) они деформируются
таким образом, что смещают
тропомиозиновую нить вглубь желобка
между актиновыми мономерами, в
результате чего контакт миозина с
актином становится возможным. |
- 2007 -